图1 主成分分析
注册
登录
English Version
陕西省科技期刊编辑学会

您当前的位置:
首页 >
文章列表页 >
饱和烷烃处理下紫花苜蓿(Medicago sativa )的转录学特征
西北大学生物学科办学100周年专刊 | 更新时间:2024-09-30
|
饱和烷烃处理下紫花苜蓿(
Medicago sativa )的转录学特征Analysis of transcriptome data of
Medicago sativa treated with saturated alkanes- 西北大学学报(自然科学版) 2024年54卷第5期 页码:889-898
DOI:10.16152/j.cnki.xdxbzr.2024-05-011
中图分类号: S541纸质出版日期:2024-10-25,
收稿日期:2024-07-30
移动端阅览

引用本文
阅读全文PDF
摘要
为探究紫花苜蓿在石油污染下的耐受机理,采用超声碎促溶的方法,将3种有机物(十二烷、十六烷和二十四烷)配置成质量分数均为1%的混合溶液,模拟饱和烷烃污染对紫花苜蓿幼苗进行处理,分别对污染0,6, 24 h的植株取样进行转录组学分析,共获得1 431个差异表达基因(DEGs)。GO富集分析表明,这些DEGs主要涉及蛋白结合、代谢途径和催化活性等;KEGG富集分析表明,DEGs主要富集到植物病原体相互作用、MAPK信号通路和光合生物碳固定途径等。qRT-PCR验证转录组结果可靠。研究结果为研究植物降解和耐受原油中饱和石油烃污染机制原理及后续筛选和培育耐石油污染植物提供理论依据。
译
Abstract
To explore the tolerance mechanism of Medicago sativa under oil pollution, this study adopted the method of ultrasonic crushing and solubilization to prepare a mixed solution of three organic compounds (Dodecane, n-Hexadecane, and n-Tetracosane) at a concentration of 1% to simulate saturated alkanes and treat Medicago sativa seedlings. Samples were taken from plants exposed to pollution for 0h, 6h, and 24h for transcriptomic analysis. The results showed that a total of 1431 DEGs were obtained. GO enrichment analysis indicated that these DEGs were mainly involved in protein binding, metabolic pathways, and catalytic activity. KEGG enrichment analysis showed that DEGs were mainly enriched in plant pathogen interaction, MAPK signaling pathway and photosynthetic biological carbon fixation pathway. qRT-PCR was further used to verify the reliability of the results. This study provides theoretical basis for exploring the mechanism of plant degradation and tolerance of saturated petroleum hydrocarbon in crude oil pollution, as well as subsequent screening and cultivation of petroleum-tolerant plants through transcriptome analysis of Medicago sativa treated with saturated alkanes.
译
石油由气态、液态和固态的烃类组成的混合物,通常贮藏在地壳上层部分[
译
紫花苜蓿(Medicago sativa)属于豆科苜蓿属,为多年生草本植物,主要种植在中国北方干旱、半干旱地区[
译
紫花苜蓿作为一种重要的牧草,其研究已经越来越深入。王天楚等将紫花苜蓿的种子在不同浓度的镉溶液中处理后继续培养生长,并对其幼苗进行指标测定,发现紫花苜蓿对于镉具有一定的富集作用[
译
1 结果
1.1 紫花苜蓿RNA测序质量
对饱和烷烃胁迫处理前(0 h)和处理后6 h和24 h的紫花苜蓿植株进行转录组的高通量测序。各个样品的GC质量分数均超过42%,且Q20>96%,Q30>94%(见
译
表1
转录组数据质控分析
Tab.1
Transcriptome data quality control and analysis
| 样本信息 | 有效数据 | Q20/% | Q30/% | GC质量分数/% |
|---|---|---|---|---|
| CK-1 | 51137498 | 97.03 | 94.88 | 43.14 |
| CK-2 | 44552210 | 96.83 | 94.49 | 42.93 |
| CK-3 | 43676428 | 96.93 | 94.64 | 43.22 |
| WR-6 h-1 | 47270614 | 96.93 | 94.67 | 43.20 |
| WR-6 h-2 | 42207482 | 96.86 | 94.52 | 43.09 |
| WR-6 h-3 | 47222008 | 96.97 | 94.72 | 42.97 |
| WR-24 h-1 | 48684712 | 96.87 | 94.54 | 42.90 |
| WR-24 h-2 | 45004252 | 96.82 | 94.59 | 42.70 |
| WR-24 h-3 | 45496924 | 96.75 | 94.47 | 42.72 |
1.2 主成分(Principal Component Analysis)分析
为了降低数据的复杂性,评估各个样品间的重复性,对9个测序样品进行了主成分分析。结果显示,对照组和处理组之间存在差异,且对照组CK-1、CK-2和CK-3之间重复性较好,模拟石油污染胁迫6 h的WR-6h-1、WR-6h-2和WR-6h-3较为集中,模拟石油污染胁迫24 h的WR-24h-1、WR-24h-2和WR-24h-3的距离较近,样品间重复性合理(见
译
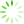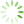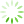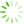
Fig.1 Principal component analysis
注: PC1为主成分1对区分样本的贡献度,PC2为主成分2对区分样本的贡献度。
1.3 差异表达基因分析
使用基于负二项分布的DESeq2软件对Raw Counts进行分析,并将Padjust<0.05和|log2FC|≥1作为差异基因的筛选标准。结果显示,与对照(CK)组相比,在模拟石油污染胁迫6 h后,共检测到783个差异表达基因,其中480个基因表达上调,而303个基因表达下调。当模拟石油污染胁迫时间达到24 h时,差异表达基因的数量变为582个,其中434个基因表达上调,148个基因表达下调(见
译
图2 基因差异分析火山图
Fig.2 Volcano plot of gene differential expression analysis
注: FC (Fold Change)为差异倍数。
Venn图展示了各个基因集中基因的个数及它们间的重叠关系。将3组CKvsWR-6h、CKvsWR-24h和WR-6hvsWR-24h的DEGs进行比较得到
译
图3 差异基因表达 Venn 图
Fig.3 Venn diagram of differential gene expression
1.4 差异表达基因的GO富集分析
GO数据库(Gene Ontology)是由基因本体联合会(Gene Ontology Consortium)所建立的,是一个国际标准化的基因功能分类体系,可以对基因和蛋白的功能进行限定和描述。分别将CKvsWR-6h、CKvsWR-24h和WR-6hvsWR-24h的DCGs进行GO功能富集分析。
译
结果显示,在CKvsWR-6h组〔见
译
图4 转录组差异表达基因的 GO富集分析
Fig.4 GO enrichment analysis of differentially expressed genes from transcriptome data
在CKvsWR-24h组〔见
译
在WR-6hvsWR-24h组〔见
译
以上结果表明,各类膜结构在响应烷烃胁迫时发挥重要的作用,水杨酸、谷胱甘肽代谢过程在模拟石油污染下紫花苜蓿的响应方面产生一定影响,通过调节氧化还原系统及抗性信号转导帮助其缓解烷烃污染胁迫给机体带来的损伤。
译
1.5 差异表达基因的KEGG富集分析
KEGG (Kyoto Encyclopedia of Genes and Genomes)数据库是一个整合了基因组、化学和系统功能信息的综合性数据库,包含了基因、蛋白质、代谢物和信号通路等生物分子的综合信息。将CKvsWR-6h、CKvsWR-24h和 WR-6hvsWR-24h的差异表达基因进行了KEGG富集分析。
译
结果显示,在CKvsWR-6h组〔见
译
图5 转录组差异表达基因的KEGG富集分析
Fig.5 KEGG enrichment analysis of differentially expressed genes in transcriptome profiling
1.6 差异表达基因的qRT-PCR验证
转录组测序共获得了1 431个差异表达基因,选取在抗氧化相关信号通路、非生物胁迫相关的信号通路和生长发育相关的信号通路中共10个最显著差异表达的基因进行验证。qRT-PCR验证结果如
译
图6 qRT-PCR验证差异表达基因
Fig.6 Validation of differentially expressed genes by qRT-PCR
注: p值,也称显著性值。以0 h为对照,*p<0.05,**p<0.01和***p<0.001。
2 讨论与结论
紫花苜蓿作为一种重要的牧草,其生长和发育过程中常会受到各种环境胁迫的影响,这些胁迫可能来自干旱[
译
本研究对模拟石油污染处理不同时间的紫花苜蓿植株的转录组进行测序,进而研究紫花苜蓿应对模拟石油污染胁迫的深层机制,对于土壤修复和生态环境保护具有重要意义。本研究从CK vsWR-6h和CK vsWR-24 h中分别获得783个和582个差异表达基因〔见
译
RT-qPCR具有高灵敏度、高特异性、高通量、实时监测且不受物种限制[
译
3 材料与方法
3.1 植物材料
以紫花苜蓿作为研究材料,用dH2O浸泡过夜后播种到营养土和蛭石1∶1配比的混合土中,在人工气候室(光照强度为100 μmol·m-2;温度25 ℃;光周期为16 h光照/8 h黑暗;空气湿度 65%)内进行培养。
译
3.2 模拟石油污染下的胁迫处理
待播种一周之后,将紫花苜蓿幼苗从土壤里移出,用清水洗干净其根部并移栽到水培箱中进行水培,加入适量的Hoagland培养液[
译
1%的模拟饱和烷烃污染混合溶液的配置方法如下:分别称取1 g的十二烷、十六烷和二十四烷于97 mL的ddH2O中,使用超声仪频率为45 kHZ超声30 min直至形成乳浊液。
译
分别收集胁迫6 h和24 h后的紫花苜蓿植株,以胁迫前(0 h)的紫花苜蓿植株作为对照,采集的紫花苜蓿植株经液氮研磨成粉末后立即放入-80 ℃冰箱中保存。每个处理组重复3次,每个样品共收集1 g材料。
译
3.3 转录组测序
所有紫花苜蓿样本基于Illumina Novaseq 6000 测序平台完成转录组的测序,测序采用Illumina TruseqTM RNA sample prep Kit方法进行文库构建。
译
为保证后续生物信息分析的准确性,使用fastp软件对原始测序数据进行过滤,从而得到高质量的测序数据(clean data),以保证后续分析的顺利进行。使用HISAT2[
译
3.4 差异表达基因DEGs的富集分析
使用软件RSEM[
译
3.5 基因的qRT-PCR验证
将选取的10个显著差异基因进行qRT-PCR分析以验证数据的可靠性。使用Trizol法提取对照组和处理组样品的总RNA,cDNA按照PrimeScript 1st Strand cDNA试剂盒说明书反转录合成。以GAPDH作为内参基因,各基因序列引物设计在附录中(见
译
表2
qRT-PCR引物序列
Tab.2
qRT-PCR primer sequences
| 基因编号 | 基因名称 | 正向引物 | 反向引物 |
|---|---|---|---|
| MsG0680034725.01 | protein REDOX2-Like | TTGGCAGCAAGGGAAACTGA | TCCAAGAGCTGACCAAGCAC |
| MsG0580024680.01 | NAC family transcription factor | TGCAATGTCCCTCCTATGGC | AGGGGAGATCCAGAAGGTTGA |
| MsG0880044306.01 | gibberellin 2-beta-dioxygenase 1 | TGCACCTTTGCCTTGTCTCA | TCTCTCAAAGTGACAAAGCCT |
| MsG0380015231.01 | glutathione S-transferase,amino-terminal | ACACAAGGCATAGCATAGCATA | GTCCACCAAAACATAGTGGGG |
| MsG0780039504.01 | cytochrome P450 76T24 | GAGCCGCAATGAAATGGTGC | ATAGCTCTGCCATGACCCAC |
| MsG0480021753.01 | L-ascorbate oxidase homolog | GCACAAGTGAATGGCAAGCA | CTTCAGAGGAGTGTCGGCTG |
| MsG0380016748.01 | peroxidase P7 | AGAAACTCTGCTCGTGGATTTG | CAGCACATGATACAACGCCAG |
| MsG0280008006.01 | flowering-promoting factor 1-like proten 1 | CAATCACGGCATCAGCGAGGAG | CTTGCGTAGCCAACACTCTCACC |
| MsG0580028960.01 | transcription factor MYB1 | AGGTGTTGGTTCATACCGCA | TGAGCGTTTGTGGAACTGGA |
| MsG0180000525.01 | transcription factor WRKY7 | GACCAACATGTGTGAGAACTATGA | ACTTTCACTCTCAGCTTTTCTCTTT |
| * | GAPDH | TCATTCCGTGTCCCAACCG | CCACATCATCTTCAGTGTAACCCA |
参考文献
[1]
KULYUKHIN S A, KOMAROV V B, SELIVERSTOV A, et al. Assessment of the applicability of an iron-containing composite for the purification of transformer oils[J]. Theoretical Foundations of Chemical Engineering, 2020, 54(5): 1057-1060. [百度学术]
[2]
HUSSAIN F, KHAN A H, HUSSAIN I, et al. Soil conditioners improve rhizodegradation of aged petroleum hydrocarbons and enhance the growth of Lolium multiflorum[J]. Environmental Science and Pollution Research, 2021, 29: 9097-9109 [百度学术]
[3]
胡玲君. 植物修复技术在土壤污染治理中的应用[J]. 黑龙江环境通报, 2022, 35(2): 80-81. [百度学术]
HU L J. Application of phytoremediation technology in soil pollution control[J]. Heilongjiang Environmental Bulletin, 2022, 35(2): 80-81. [百度学术]
[4]
HOU J, WANG Q, LIU W, et al. Soil microbial community and association network shift induced by several tall fescue cultivars during the phytoremediation of a petroleum hydrocarbon-contaminated soil[J]. Science of the Total Environment, 2021, 792: 148411. [百度学术]
[5]
W<inline-formula><alternatives><graphic specific-use="print" xlink:href="media/1000-274X-54-05-011-I001.jpg"/><graphic specific-use="big" xlink:href="alternativeImage/1000-274X-54-05-011-I001.jpg"/><graphic specific-use="small" xlink:href="alternativeImage/1000-274X-54-05-011-I001c.jpg"/></alternatives></inline-formula>ÓKA D, PLACEK A, SMOL M, et al. The efficiency and economic aspects of phytoremediation technology using Phalaris arundinacea L. and Brassica napus L. combined with compost and nano SiO2 fertilization for the removal of PAH’s from soil[J]. Journal of Enrironmental Management, 2019, 234(15): 311-319. [百度学术]
[6]
徐艳, 史高琦. 王曙光,等.紫花苜蓿对石油污染土壤的修复响应[J]. 环境科学与技术, 2018, 41(增刊2): 25-28. [百度学术]
XU Y, SHI G Q, WANG S G, et al. Remediation response of Medicago sativa to oil contaminated soil[J]. Environmental Science and Technology, 2018, 41(S2): 25-28. [百度学术]
[7]
CAI B, MA J, YAN G X, et al. Comparison of phytoremediation, bioaugmentation and natural attenuation for remediating saline soil contaminated by heavy crude oil [J]. Biochemical Engineering Journal, 2016, 112: 170-177. [百度学术]
[8]
孟楠, 王萌, 陈莉, 等. 不同草本植物间作对Cd污染土壤的修复效果[J]. 中国环境科学, 2018, 38(7): 2618-2624. [百度学术]
MENG N, WANG M, CHEN L, et al. Remediation effect of different herbaceous plants intercropping on Cd contaminated soil[J]. China Environmental Science, 2018, 38(7): 2618-2624. [百度学术]
[9]
LIU W X, HOU J Y, WANG Q L, et al. Collection and analysis of rootexudates of Festuca arundinacea L. and their role in facilitating the phytoremediation of petroleum-contaminated soil [J]. Plant and Soil, 2015, 389(1/2): 109-119. [百度学术]
[10]
王运涛, 孟德斌, 于林清, 等. 8个紫花苜蓿材料在呼和浩特地区的抗寒性和生产性能比较[J]. 中国草地学报, 2022, 44(6): 60-66. [百度学术]
WANG Y T, MENG D B, YU L Q, et al. Comparison of cold resistance and production performance of eight Medicago sativa materials in Hohhot Area [J]. Chinese Journal of Grassland, 2022, 44(6): 60-66. [百度学术]
[11]
孟宣辰, 马鹏程, 马杰, 等. 黄土高原半干旱区紫花苜蓿覆膜垄沟和施肥效应研究[J]. 草地学报, 2021, 14(6): 5-7. [百度学术]
MENG X C, MA P C, MA J, et al. Effects of mulching ridge and furrow and fertilization on alfalfa in the semi-arid region of the Loess Plateau[J]. Journal of Grassland, 2021, 14(6): 5-7. [百度学术]
[12]
边佳辉, 刘白扬, 李宗英, 等. 紫花苜蓿基因工程改良研究进展[J]. 草业科学, 2020, 37(1): 139-155. [百度学术]
BIAN J H, LIU B Y, LI Z Y, et al. Research progress on genetic engineering improvement of alfalfa[J]. Pratacultural Science, 2020, 37(1): 139-155. [百度学术]
[13]
SINGER S D, HANNOUFA A, ACHARYA S. Molecular improvement of alfalfa for enhanced productivity and adaptability in a changing environment[J]. Plant Cell Environ, 2018, 41(9): 1955-1971. [百度学术]
[14]
李会彬, 王丽宏, 边秀举, 等. 河北省紫花苜蓿农药残留状况调查与分析[J]. 河北农业科学, 2020, 24(1): 61-64. [百度学术]
LI H B, WANG L H, BIAN X J, et al. Investigation and analysis of pesticide residues in Medicago sativa in Hebei Province [J]. Journal of Hebei Agricultural Sciences, 2020, 24(1): 61-64. [百度学术]
[15]
CHEN L, BEIYUAN J, HU W, et al. Phytoremediation of potentially toxic elements (PTEs) contaminated soils using alfalfa (Medicago sativa L.): A comprehensive review[J]. Chemosphere, 2022, 293: 133577. [百度学术]
[16]
ZHANG F B, HUANG L H, YONG M Y, et al. Rainfall simulation experiments indicate that biochar addition enhances erosion of loess-derived soils[J]. Land Degradation & Development, 2019, 30: 2272-2286. [百度学术]
[17]
王天楚, 段士鑫, 吴怀欣, 等. 基于相关分析和主成分分析的紫花苜蓿抗镉胁迫研究[J]. 南方农业学报, 2023, 54(10): 2878-2887. [百度学术]
WANG T C, DUAN S X, WU H X, et al. Study on cadmium stress resistance of alfalfa based on correlation analysis and principal component analysis [J]. Journal of Southern Agriculture, 2023, 54(10): 2878-2887. [百度学术]
[18]
熊军波, 杨青川, 蔡化, 等. 紫花苜蓿根响应盐胁迫的比较蛋白质组学分析[J]. 湖北农业科学, 2015, 54(21): 5422-5428. [百度学术]
XIONG J B, YANG Q C, CAI H, et al. Comparative proteomics analysis of alfalfa roots in response to salt stress [J]. Hubei Agricultural Sciences, 2015, 54(21): 5422-5428. [百度学术]
[19]
GAO Q, YU R, MA X, et al. Transcriptome analysis for salt-responsive genes in two different alfalfa (Medicago sativa L.) cultivars and functional analysis of MsHPCA1[J]. Plants (Basel), 2024, 13(8): 1073-1091. [百度学术]
[20]
ZHANG C, SHI S. Physiological and proteomic responses of contrasting alfalfa (Medicago sativa L.) varieties to PEG-induced osmotic stress[J]. Front Plant Science, 2018, 9: 753011. [百度学术]
[21]
于浩然, 贾玉山, 刘鹰昊, 等. 土壤盐碱度和留茬高度对苜蓿农艺性状及干草品质的影响[J]. 西北农林科技大学学报(自然科学版), 2020, 48(1): 33-39. [百度学术]
YU H R, JIA Y S, LIU Y H, et al. Effects of soil salinity and stubble height on agronomic traits and hay quality of alfalfa [J]. Journal of Northwest A&F University (Natural Science Edition), 2020, 48(1): 33-39. [百度学术]
[22]
闫志强, 陈银萍, 蘧苗苗, 等. 镉胁迫对紫花苜蓿幼苗生理特性和镉富集的影响[J]. 广西植物, 2019, 39(2): 218-227. [百度学术]
YAN Z Q, CHEN Y P, QU M M, et al. Effects of cadmium stress on physiological characteristics and cadmium enrichment in alfalfa seedlings [J]. Guihaia, 2019, 39(2): 218-227. [百度学术]
[23]
曹婧, 李向林, 万里强. 锰胁迫对紫花苜蓿生理和生长特性的影响[J]. 中国草地学报, 2019, 41(6): 15-22. [百度学术]
CAO J, LI X L, WAN L Q. Effects of manganese stress on physiological and growth characteristics of alfalfa[J]. Chinese Journal of Grassland, 2019, 41(6): 15-22. [百度学术]
[24]
赵颖. 基于转录组分析揭示一氧化氮调控紫花苜蓿响应干旱胁迫的机理[D]. 兰州: 甘肃农业大学, 2020. [百度学术]
[25]
LUO D, ZHOU Q, WU Y G, et al. Full-length transcript sequencing and comparative transcriptomic analysis to evaluate the contribution of osmotic and ionic stress components towards salinity tolerance in the roots of cultivated alfalfa (Medicago sativa L.)[J]. BMC Plant Biology, 2019, 19(1): 32: 1-20. [百度学术]
[26]
WANG Z, GERSTEIN M, SNYDER M. RNA-Seq: A revolutionary tool for transcriptomics.[J]. Nature Reviews Genetics, 2009, 10(1): 57-63. [百度学术]
[27]
黄申, 闫茗熠, 陈梦月, 等. 基于转录组测序和RT-qPCR技术的烟草糖酯合成基因挖掘[J]. 轻工学报, 2023, 38(6): 78-84. [百度学术]
HUANG S, YAN M Y, CHEN M Y, et al. Mining of tobacco sugar ester synthesis genes based on transcriptome sequencing and RT-qPCR technology [J]. Journal of Light Industry, 2023, 38(6): 78-84. [百度学术]
[28]
魏春梅, 栾威, 代娅, 等. 比较转录组研究钛离子对紫花苜蓿基因表达的影响[J]. 应用与环境生物学报, 2019, 25(1): 117-127. [百度学术]
WEI C M, LUAN W, DAI Y, et al. Comparative transcriptome study on the effects of titanium ions on gene expression in alfalfa [J]. Chinese Journal of Applied and Environmental Biology, 2019, 25(1): 117-127. [百度学术]
[29]
YAN H F, MA G H, JAIME A, et al. Genome-wide identification and analysis of NAC transcription factor family in twodiploid wild relatives of cultivated sweet potato uncovers potentia lNAC genes related to drought tolerance[J]. Front Genet, 2021, 12: 744220. [百度学术]
[30]
JU Y, YUE X, MIN Z, et al. VvNAC17, a novel stress-responsive grapevine (Vitis vinifera L.) NAC transcription factor, increases sensitivity to abscisic acid and enhances salinity, freezing, and drought tolerance in transgenic Arabidopsis[J]. Plant Physiology and Biochemistry, 2020, 146: 98-111. [百度学术]
[31]
李晓宇, 李文颖, 杨成超. 镉胁迫下中辽1号杨转录组分析[J]. 西北农林科技大学学报(自然科学版), 2024, 52(6): 29-39. [百度学术]
LI X Y, LI W Y, YANG C C. Transcriptome analysis of Zhongliao No.1 poplar under cadmium stress [J]. Journal of Northwest A&F University (Natural Science Edition), 2024, 52(6): 29-39. [百度学术]
[32]
FENG X, XU Y, PENG L, et al. TaEXPB7-B, a β-expansin gene involved in low-temperature stress and abscisic acid responses, promotes growth and cold resistance in Arabidopsis thaliana[J]. Journal of Plant Physiology, 2019, 240: 153004. [百度学术]
[33]
陆程张. Na2CO3胁迫下玉米苗期根部转录组表达谱分析[D]. 延吉: 延边大学, 2021. [百度学术]
[34]
毕博远, 韩凤朋. 黄土高原不同种植年限紫花苜蓿根系分泌物GC-MS分析[J]. 草地学报, 2018, 26(3): 611-617. [百度学术]
BI B Y, HAN F P. GC-MS analysis of root exudates from alfalfa planted in the Loess Plateau for different years[J]. Acta Agrestia Sinica, 2018, 26(3): 611-617. [百度学术]
[35]
KIM D, LANGMEAD B, SALZBERG S L. HISAT: A fast spliced aligner with low memory requirements[J]. Nature Methods, 2015, 12(4): 357-360. [百度学术]
[36]
TRAPNELL C, WILLIAMS B A, PERTEA G, et al. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation[J]. Nature Biotechnology, 2010, 28(5): 511-515. [百度学术]
[37]
LI B, DEWEY C N. RSEM: Accurate transcript quantification from RNA-Seq data with or without a reference genome[J]. BMC Bioinformatics, 2011, 12(1): 1-16. [百度学术]
[38]
朱艳霞, 姜建萍, 郭晓云, 等. 药用植物青葙全长转录组测序分析 [J]. 中国现代中药, 2023, 25(7): 1463-1472. [百度学术]
ZHU Y X, JIANG J P, GUO X Y, et al. Full-length transcriptome sequencing analysis of the medicinal plant Celosia argentea [J]. Modern Chinese Medicine, 2023, 25(7): 1463-1472. [百度学术]
[39]
MENG A, WEI S, YAN L, et al. Bioniformatic analysis and validation of ardiac hypertrophy-related genes [J]. Zenodo, 2023, 42(2): 159-167. [百度学术]
[40]
LIU L, LIU D, WANG Z, et al. Exogenous allantoin improves the salt tolerance of sugar beet by increasing putrescine metabolism and antioxidant activities[J]. Plant Physiology and Biochemistry, 2020, 154: 699-713. [百度学术]
151
浏览量
28
下载量
0
CSCD
文章被引用时,请邮件提醒。
提交
工具集
下载
参考文献导出
分享
收藏
添加至我的专辑
相关文章
暂无数据
相关作者
暂无数据
相关机构
暂无数据
- 地址:陕西省西安市太白北路229号
- 联系电话:029-88302822 Email:xdxbzr@nwu.edu.cn
- 技术支持由北京北大方正电子有限公司提供 陕ICP备05010980号-4
 京公网安备11010802024621
京公网安备11010802024621 - 本系统建议在Chrome、 IE9+ 以上版本浏览器阅读本站内容,360浏览器请切换至极速模式
- Cookies帮助我们提供服务并提供个性化体验。使用本网站,即表示您同意我们使用Cookies
0